Project FOMC19244_48S_Ahmed services include NGS sequencing of the V1V3 region of the 16S rRNA gene amplicons from the samples. First and foremost, please
download this report, as well as the sequence raw data from the download links provided below.
These links will expire after 60 days. We cannot guarantee the availability of your data after 60 days.
Full Bioinformatics analysis service was requested. We provide many analyses, starting from the raw sequence quality and noise filtering, pair reads merging, as well as chimera filtering for the sequences, using the
DADA2 denosing algorithm and pipeline.
We also provide many downstream analyses such as taxonomy assignment, alpha and beta diversity analyses, and differential abundance analysis.
For taxonomy assignment, most informative would be the taxonomy barplots. We provide an interactive barplots to show the relative abundance of microbes at different taxonomy levels (from Phylum to species) that you can choose.
If you specify which groups of samples you want to compare for differential abundance, we provide both ANCOM and LEfSe differential abundance analysis.
The samples were processed and analyzed with the ZymoBIOMICS® Service: Targeted
Metagenomic Sequencing (Zymo Research, Irvine, CA).
DNA Extraction: If DNA extraction was performed, the following DNA
extraction kit was used according to the manufacturer’s instructions:
☑
ZymoBIOMICS®-96 MagBead DNA Kit (Zymo Research, Irvine, CA)
☐
N/A (DNA Extraction Not Performed)
Elution Volume: 50µL
Additional Notes: NA
Targeted Library Preparation: The DNA samples were prepared for targeted
sequencing with the Quick-16S™ NGS Library Prep Kit (Zymo Research, Irvine, CA).
These primers were custom designed by Zymo Research to provide the best coverage
of the 16S gene while maintaining high sensitivity. The primer sets used in this project
are marked below:
☐
Quick-16S™ Primer Set V1-V2 (Zymo Research, Irvine, CA)
☑
Quick-16S™ Primer Set V1-V3 (Zymo Research, Irvine, CA)
☐
Quick-16S™ Primer Set V3-V4 (Zymo Research, Irvine, CA)
☐
Quick-16S™ Primer Set V4 (Zymo Research, Irvine, CA)
☐
Quick-16S™ Primer Set V6-V8 (Zymo Research, Irvine, CA)
Additional Notes: NA
The sequencing library was prepared using an innovative library preparation process in
which PCR reactions were performed in real-time PCR machines to control cycles and
therefore limit PCR chimera formation. The final PCR products were quantified with
qPCR fluorescence readings and pooled together based on equal molarity. The final
pooled library was cleaned up with the Select-a-Size DNA Clean & Concentrator™
(Zymo Research, Irvine, CA), then quantified with TapeStation® (Agilent Technologies,
Santa Clara, CA) and Qubit® (Thermo Fisher Scientific, Waltham, WA).
Control Samples: The ZymoBIOMICS® Microbial Community Standard (Zymo
Research, Irvine, CA) was used as a positive control for each DNA extraction, if
performed. The ZymoBIOMICS® Microbial Community DNA Standard (Zymo Research,
Irvine, CA) was used as a positive control for each targeted library preparation.
Negative controls (i.e. blank extraction control, blank library preparation control) were
included to assess the level of bioburden carried by the wet-lab process.
Sequencing: The final library was sequenced on Illumina® NextSeq 2000™ with a p1
(Illumina, Sand Diego, CA) reagent kit (600 cycles). The sequencing was performed
with 25% PhiX spike-in.
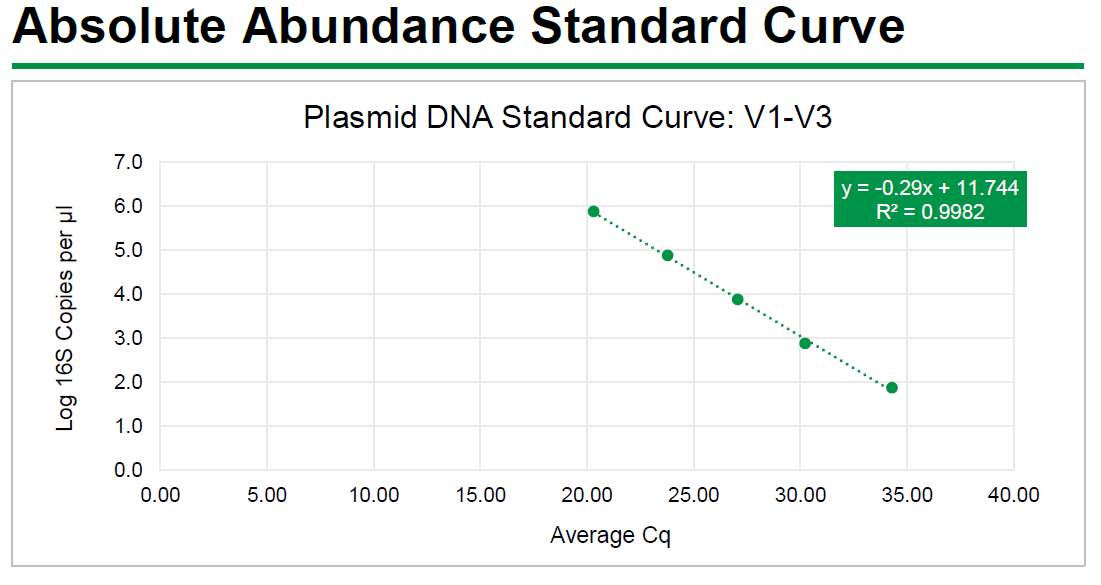
Absolute Abundance Quantification*: A quantitative real-time PCR was set up with a
standard curve. The standard curve was made with plasmid DNA containing one copy
of the 16S gene and one copy of the fungal ITS2 region prepared in 10-fold serial
dilutions. The primers used were the same as those used in Targeted Library
Preparation. The equation generated by the plasmid DNA standard curve was used to
calculate the number of gene copies in the reaction for each sample. The PCR input
volume (2 µl) was used to calculate the number of gene copies per microliter in each
DNA sample.
The number of genome copies per microliter DNA sample was calculated by dividing
the gene copy number by an assumed number of gene copies per genome. The value
used for 16S copies per genome is 4. The value used for ITS copies per genome is 200.
The amount of DNA per microliter DNA sample was calculated using an assumed
genome size of 4.64 x 106 bp, the genome size of Escherichia coli, for 16S samples, or
an assumed genome size of 1.20 x 107 bp, the genome size of Saccharomyces
cerevisiae, for ITS samples. This calculation is shown below:
Calculated Total DNA = Calculated Total Genome Copies × Assumed Genome Size (4.64 × 106 bp) ×
Average Molecular Weight of a DNA bp (660 g/mole/bp) ÷ Avogadro’s Number (6.022 x 1023/mole)
* Absolute Abundance Quantification is only available for 16S and ITS analyses.
The absolute abundance standard curve data can be viewed in Excel here:
The absolute abundance standard curve is shown below:
The complete report of your project, including all links in this report, can be downloaded by clicking the link provided below. The downloaded file is a compressed ZIP file and once unzipped, open the file “REPORT.html” (may only shown as "REPORT" in your computer) by double clicking it. Your default web browser will open it and you will see the exact content of this report.
Please download and save the file to your computer storage device. The download link will expire after 60 days upon your receiving of this report.
Complete report download link:
To view the report, please follow the following steps:
1.
Download the .zip file from the report link above.
2.
Extract all the contents of the downloaded .zip file to your desktop.
3.
Open the extracted folder and find the "REPORT.html" (may shown as only "REPORT").
4.
Open (double-clicking) the REPORT.html file. Your default browser will open the top age of the complete report. Within the
report, there are links to view all the analyses performed for the project.
The raw NGS sequence data is available for download with the link provided below. The data is a compressed ZIP file and can be unzipped to individual sequence files.
Since this is a pair-end sequencing, each of your samples is represented by two sequence files, one for READ 1,
with the file extension “*_R1.fastq.gz”, another READ 2, with the file extension “*_R1.fastq.gz”.
The files are in FASTQ format and are compressed. FASTQ format is a text-based data format for storing both a biological sequence
and its corresponding quality scores. Most sequence analysis software will be able to open them.
The Sample IDs associated with the R1 and R2 fastq files are listed in the table below:
Sample ID
Original Sample ID
Read 1 File Name
Read 2 File Name
F19244.S10
original sample ID here
zr19244_10V1V3_R1.fastq.gz
zr19244_10V1V3_R2.fastq.gz
F19244.S11
original sample ID here
zr19244_11V1V3_R1.fastq.gz
zr19244_11V1V3_R2.fastq.gz
F19244.S12
original sample ID here
zr19244_12V1V3_R1.fastq.gz
zr19244_12V1V3_R2.fastq.gz
F19244.S13
original sample ID here
zr19244_13V1V3_R1.fastq.gz
zr19244_13V1V3_R2.fastq.gz
F19244.S14
original sample ID here
zr19244_14V1V3_R1.fastq.gz
zr19244_14V1V3_R2.fastq.gz
F19244.S15
original sample ID here
zr19244_15V1V3_R1.fastq.gz
zr19244_15V1V3_R2.fastq.gz
F19244.S16
original sample ID here
zr19244_16V1V3_R1.fastq.gz
zr19244_16V1V3_R2.fastq.gz
F19244.S17
original sample ID here
zr19244_17V1V3_R1.fastq.gz
zr19244_17V1V3_R2.fastq.gz
F19244.S18
original sample ID here
zr19244_18V1V3_R1.fastq.gz
zr19244_18V1V3_R2.fastq.gz
F19244.S19
original sample ID here
zr19244_19V1V3_R1.fastq.gz
zr19244_19V1V3_R2.fastq.gz
F19244.S01
original sample ID here
zr19244_1V1V3_R1.fastq.gz
zr19244_1V1V3_R2.fastq.gz
F19244.S20
original sample ID here
zr19244_20V1V3_R1.fastq.gz
zr19244_20V1V3_R2.fastq.gz
F19244.S21
original sample ID here
zr19244_21V1V3_R1.fastq.gz
zr19244_21V1V3_R2.fastq.gz
F19244.S22
original sample ID here
zr19244_22V1V3_R1.fastq.gz
zr19244_22V1V3_R2.fastq.gz
F19244.S23
original sample ID here
zr19244_23V1V3_R1.fastq.gz
zr19244_23V1V3_R2.fastq.gz
F19244.S24
original sample ID here
zr19244_24V1V3_R1.fastq.gz
zr19244_24V1V3_R2.fastq.gz
F19244.S25
original sample ID here
zr19244_25V1V3_R1.fastq.gz
zr19244_25V1V3_R2.fastq.gz
F19244.S26
original sample ID here
zr19244_26V1V3_R1.fastq.gz
zr19244_26V1V3_R2.fastq.gz
F19244.S27
original sample ID here
zr19244_27V1V3_R1.fastq.gz
zr19244_27V1V3_R2.fastq.gz
F19244.S28
original sample ID here
zr19244_28V1V3_R1.fastq.gz
zr19244_28V1V3_R2.fastq.gz
F19244.S29
original sample ID here
zr19244_29V1V3_R1.fastq.gz
zr19244_29V1V3_R2.fastq.gz
F19244.S02
original sample ID here
zr19244_2V1V3_R1.fastq.gz
zr19244_2V1V3_R2.fastq.gz
F19244.S30
original sample ID here
zr19244_30V1V3_R1.fastq.gz
zr19244_30V1V3_R2.fastq.gz
F19244.S31
original sample ID here
zr19244_31V1V3_R1.fastq.gz
zr19244_31V1V3_R2.fastq.gz
F19244.S32
original sample ID here
zr19244_32V1V3_R1.fastq.gz
zr19244_32V1V3_R2.fastq.gz
F19244.S33
original sample ID here
zr19244_33V1V3_R1.fastq.gz
zr19244_33V1V3_R2.fastq.gz
F19244.S34
original sample ID here
zr19244_34V1V3_R1.fastq.gz
zr19244_34V1V3_R2.fastq.gz
F19244.S35
original sample ID here
zr19244_35V1V3_R1.fastq.gz
zr19244_35V1V3_R2.fastq.gz
F19244.S36
original sample ID here
zr19244_36V1V3_R1.fastq.gz
zr19244_36V1V3_R2.fastq.gz
F19244.S37
original sample ID here
zr19244_37V1V3_R1.fastq.gz
zr19244_37V1V3_R2.fastq.gz
F19244.S38
original sample ID here
zr19244_38V1V3_R1.fastq.gz
zr19244_38V1V3_R2.fastq.gz
F19244.S39
original sample ID here
zr19244_39V1V3_R1.fastq.gz
zr19244_39V1V3_R2.fastq.gz
F19244.S03
original sample ID here
zr19244_3V1V3_R1.fastq.gz
zr19244_3V1V3_R2.fastq.gz
F19244.S40
original sample ID here
zr19244_40V1V3_R1.fastq.gz
zr19244_40V1V3_R2.fastq.gz
F19244.S41
original sample ID here
zr19244_41V1V3_R1.fastq.gz
zr19244_41V1V3_R2.fastq.gz
F19244.S42
original sample ID here
zr19244_42V1V3_R1.fastq.gz
zr19244_42V1V3_R2.fastq.gz
F19244.S43
original sample ID here
zr19244_43V1V3_R1.fastq.gz
zr19244_43V1V3_R2.fastq.gz
F19244.S44
original sample ID here
zr19244_44V1V3_R1.fastq.gz
zr19244_44V1V3_R2.fastq.gz
F19244.S45
original sample ID here
zr19244_45V1V3_R1.fastq.gz
zr19244_45V1V3_R2.fastq.gz
F19244.S46
original sample ID here
zr19244_46V1V3_R1.fastq.gz
zr19244_46V1V3_R2.fastq.gz
F19244.S47
original sample ID here
zr19244_47V1V3_R1.fastq.gz
zr19244_47V1V3_R2.fastq.gz
F19244.S48
original sample ID here
zr19244_48V1V3_R1.fastq.gz
zr19244_48V1V3_R2.fastq.gz
F19244.S04
original sample ID here
zr19244_4V1V3_R1.fastq.gz
zr19244_4V1V3_R2.fastq.gz
F19244.S05
original sample ID here
zr19244_5V1V3_R1.fastq.gz
zr19244_5V1V3_R2.fastq.gz
F19244.S06
original sample ID here
zr19244_6V1V3_R1.fastq.gz
zr19244_6V1V3_R2.fastq.gz
F19244.S07
original sample ID here
zr19244_7V1V3_R1.fastq.gz
zr19244_7V1V3_R2.fastq.gz
F19244.S08
original sample ID here
zr19244_8V1V3_R1.fastq.gz
zr19244_8V1V3_R2.fastq.gz
F19244.S09
original sample ID here
zr19244_9V1V3_R1.fastq.gz
zr19244_9V1V3_R2.fastq.gz
Please download and save the file to your computer storage device. The download link will expire after 60 days upon your receiving of this report.
DADA2 is a software package that models and corrects Illumina-sequenced amplicon errors [1].
DADA2 infers sample sequences exactly, without coarse-graining into OTUs,
and resolves differences of as little as one nucleotide. DADA2 identified more real variants
and output fewer spurious sequences than other methods.
DADA2’s advantage is that it uses more of the data. The DADA2 error model incorporates quality information,
which is ignored by all other methods after filtering. The DADA2 error model incorporates quantitative abundances,
whereas most other methods use abundance ranks if they use abundance at all.
The DADA2 error model identifies the differences between sequences, eg. A->C,
whereas other methods merely count the mismatches. DADA2 can parameterize its error model from the data itself,
rather than relying on previous datasets that may or may not reflect the PCR and sequencing protocols used in your study.
Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJ, Holmes SP. DADA2: High-resolution sample inference from Illumina amplicon data. Nat Methods. 2016 Jul;13(7):581-3. doi: 10.1038/nmeth.3869. Epub 2016 May 23. PMID: 27214047; PMCID: PMC4927377.
Analysis Procedures:
DADA2 pipeline includes several tools for read quality control, including quality filtering, trimming, denoising, pair merging and chimera filtering. Below are the major processing steps of DADA2:
Step 1. Read trimming based on sequence quality
The quality of NGS Illumina sequences often decreases toward the end of the reads.
DADA2 allows to trim off the poor quality read ends in order to improve the error
model building and pair mergicing performance.
Step 2. Learn the Error Rates
The DADA2 algorithm makes use of a parametric error model (err) and every
amplicon dataset has a different set of error rates. The learnErrors method
learns this error model from the data, by alternating estimation of the error
rates and inference of sample composition until they converge on a jointly
consistent solution. As in many machine-learning problems, the algorithm must
begin with an initial guess, for which the maximum possible error rates in
this data are used (the error rates if only the most abundant sequence is
correct and all the rest are errors).
Step 3. Infer amplicon sequence variants (ASVs) based on the error model built in previous step. This step is also called sequence "denoising".
The outcome of this step is a list of ASVs that are the equivalent of oligonucleotides.
Step 4. Merge paired reads. If the sequencing products are read pairs, DADA2 will merge the R1 and R2 ASVs into single sequences.
Merging is performed by aligning the denoised forward reads with the reverse-complement of the corresponding
denoised reverse reads, and then constructing the merged “contig” sequences.
By default, merged sequences are only output if the forward and reverse reads overlap by
at least 12 bases, and are identical to each other in the overlap region (but these conditions can be changed via function arguments).
Step 5. Remove chimera.
The core dada method corrects substitution and indel errors, but chimeras remain. Fortunately, the accuracy of sequence variants
after denoising makes identifying chimeric ASVs simpler than when dealing with fuzzy OTUs.
Chimeric sequences are identified if they can be exactly reconstructed by
combining a left-segment and a right-segment from two more abundant “parent” sequences. The frequency of chimeric sequences varies substantially
from dataset to dataset, and depends on on factors including experimental procedures and sample complexity.
Results
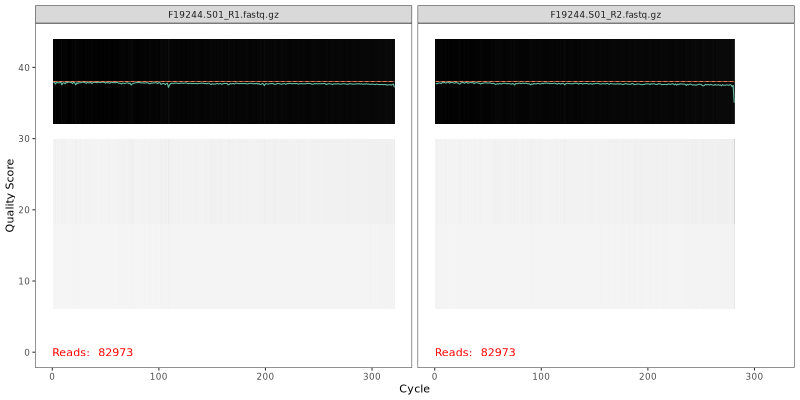
1. Read Quality Plots NGS sequence analaysis starts with visualizing the quality of the sequencing. Below are the quality plots of the first
sample for the R1 and R2 reads separately. In gray-scale is a heat map of the frequency of each quality score at each base position. The mean
quality score at each position is shown by the green line, and the quartiles of the quality score distribution by the orange lines.
The forward reads are usually of better quality. It is a common practice to trim the last few nucleotides to avoid less well-controlled errors
that can arise there. The trimming affects the downstream steps including error model building, merging and chimera calling. FOMC uses an empirical
approach to test many combinations of different trim length in order to achieve best final amplicon sequence variants (ASVs), see the next
section “Optimal trim length for ASVs”.
2. Optimal trim length for ASVs The final number of merged and chimera-filtered ASVs depends on the quality filtering (hence trimming) in the very beginning of the DADA2 pipeline.
In order to achieve highest number of ASVs, an empirical approach was used -
Create a random subset of each sample consisting of 5,000 R1 and 5,000 R2 (to reduce computation time)
Trim 10 bases at a time from the ends of both R1 and R2 up to 50 bases
For each combination of trimmed length (e.g., 300x300, 300x290, 290x290 etc), the trimmed reads are
subject to the entire DADA2 pipeline for chimera-filtered merged ASVs
The combination with highest percentage of the input reads becoming final ASVs is selected for the complete set of data
Below is the result of such operation, showing ASV percentages of total reads for all trimming combinations (1st Column = R1 lengths in bases; 1st Row = R2 lengths in bases):
R1/R2
281
271
261
251
241
231
321
81.76%
81.93%
82.16%
82.45%
82.46%
75.85%
311
81.72%
81.98%
82.22%
82.16%
76.01%
57.02%
301
81.80%
82.07%
81.93%
75.68%
57.11%
34.02%
291
81.84%
81.70%
75.31%
56.71%
34.07%
23.14%
281
81.58%
75.16%
56.54%
33.76%
23.04%
9.80%
271
75.19%
56.67%
33.74%
22.88%
9.76%
3.66%
Based on the above result, the trim length combination of R1 = 321 bases and R2 = 241 bases (highlighted red above), was chosen for generating final ASVs for all sequences.
This combination generated highest number of merged non-chimeric ASVs and was used for downstream analyses, if requested.
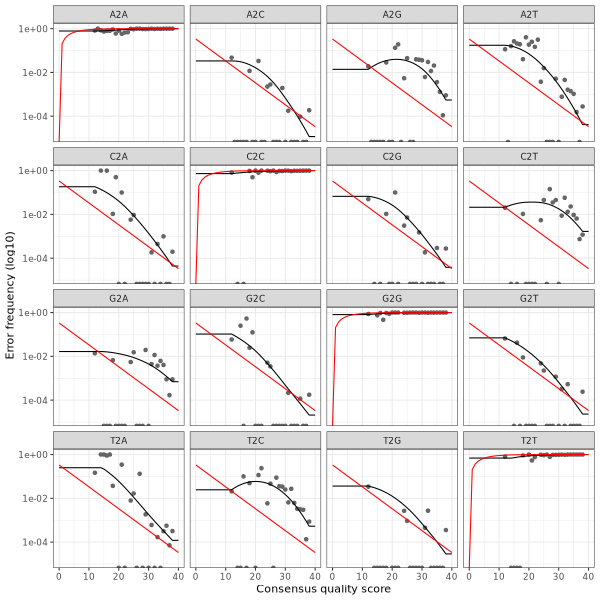
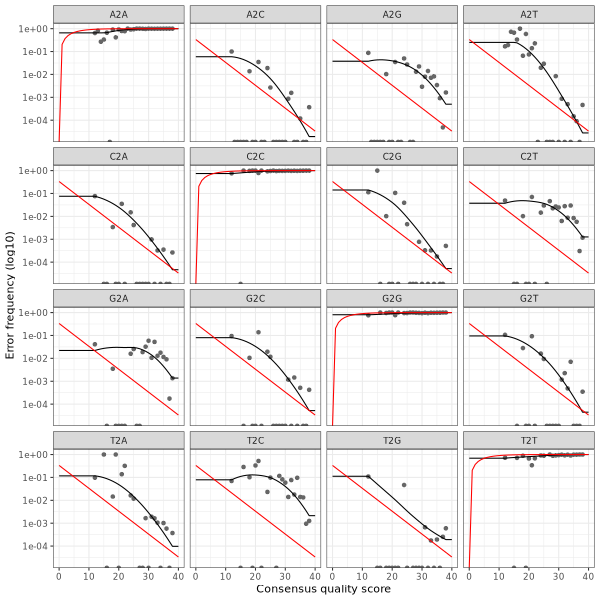
3. Error plots from learning the error rates
After DADA2 building the error model for the set of data, it is always worthwhile, as a sanity check if nothing else, to visualize the estimated error rates.
The error rates for each possible transition (A→C, A→G, …) are shown below. Points are the observed error rates for each consensus quality score.
The black line shows the estimated error rates after convergence of the machine-learning algorithm.
The red line shows the error rates expected under the nominal definition of the Q-score.
The ideal result would be the estimated error rates (black line) are a good fit to the observed rates (points), and the error rates drop
with increased quality as expected.
Forward Read R1 Error Plot
Reverse Read R2 Error Plot
The PDF version of these plots are available here:
4. DADA2 Result Summary The table below shows the summary of the DADA2 analysis,
tracking paired read counts of each samples for all the steps during DADA2 denoising process -
including end-trimming (filtered), denoising (denoisedF, denoisedF), pair merging (merged) and chimera removal (nonchim).
Sample ID
F19244.S01
F19244.S02
F19244.S03
F19244.S04
F19244.S05
F19244.S06
F19244.S07
F19244.S08
F19244.S09
F19244.S10
F19244.S11
F19244.S12
F19244.S13
F19244.S14
F19244.S15
F19244.S16
F19244.S17
F19244.S18
F19244.S19
F19244.S20
F19244.S21
F19244.S22
F19244.S23
F19244.S24
F19244.S25
F19244.S26
F19244.S27
F19244.S28
F19244.S29
F19244.S30
F19244.S31
F19244.S32
F19244.S33
F19244.S34
F19244.S35
F19244.S36
F19244.S37
F19244.S38
F19244.S39
F19244.S40
F19244.S41
F19244.S42
F19244.S43
F19244.S44
F19244.S45
F19244.S46
F19244.S47
F19244.S48
Row Sum
Percentage
input
82,973
120,857
103,275
81,012
87,639
87,539
97,033
87,960
66,123
65,193
79,144
88,006
100,012
69,994
91,117
84,811
79,047
72,552
68,839
89,444
125,663
107,703
106,631
89,197
64,072
86,349
78,945
103,483
101,730
112,372
78,860
97,783
75,190
74,828
70,343
80,436
77,094
77,419
82,486
78,902
101,625
73,130
88,898
87,682
108,654
107,807
82,578
75,853
4,198,283
100.00%
filtered
82,972
120,856
103,273
81,011
87,637
87,537
97,032
87,959
66,122
65,193
79,142
88,005
100,009
69,994
91,115
84,811
79,045
72,552
68,838
89,443
125,662
107,702
106,630
89,196
64,072
86,346
78,944
103,482
101,730
112,371
78,859
97,782
75,190
74,828
70,341
80,436
77,093
77,418
82,486
78,902
101,625
73,129
88,894
87,679
108,652
107,806
82,576
75,852
4,198,229
100.00%
denoisedF
82,195
119,717
102,246
79,932
86,936
86,871
96,293
87,461
65,558
64,839
78,413
87,426
98,831
69,171
90,078
84,168
78,152
71,675
68,193
88,852
124,710
107,031
105,620
88,485
63,283
85,488
78,128
102,911
100,633
111,461
77,721
96,565
74,594
74,164
69,780
80,085
75,988
76,300
81,808
78,260
100,683
72,597
87,962
86,531
107,653
107,087
81,580
74,919
4,159,034
99.07%
denoisedR
81,530
119,426
102,133
79,849
86,826
86,524
96,014
87,339
65,452
64,845
78,507
87,421
98,732
68,595
89,913
84,215
77,990
71,382
67,711
88,705
124,297
106,981
105,695
88,361
63,350
85,337
77,888
102,905
100,207
111,311
77,153
96,209
74,113
74,034
69,574
79,807
75,671
76,000
81,309
77,985
100,585
72,377
87,821
86,147
107,068
106,929
81,142
74,614
4,147,979
98.80%
merged
75,906
108,057
96,032
74,828
81,488
82,202
91,414
83,138
61,428
62,916
73,732
83,129
92,941
63,353
82,913
80,046
72,559
66,151
62,257
84,298
118,027
102,953
100,605
83,465
59,147
80,520
71,822
99,143
93,247
105,686
70,156
89,527
68,330
69,452
64,972
76,200
68,781
69,278
75,150
73,324
94,056
67,038
82,998
79,957
99,764
101,503
74,309
69,213
3,887,411
92.60%
nonchim
71,896
103,162
91,317
71,263
77,128
73,270
82,808
70,491
49,131
46,184
64,908
71,162
89,852
59,028
76,346
66,417
69,661
63,025
55,576
76,102
112,555
95,353
87,755
74,642
55,314
74,164
63,542
84,926
88,796
102,226
65,991
85,641
63,762
60,257
55,266
71,173
62,882
61,327
71,226
66,646
88,433
62,727
80,522
76,318
91,145
92,138
69,576
64,892
3,557,922
84.75%
This table can be downloaded as an Excel table below:
5. DADA2 Amplicon Sequence Variants (ASVs). A total of 7002 unique merged and chimera-free ASV sequences were identified, and their corresponding
read counts for each sample are available in the "ASV Read Count Table" with rows for the ASV sequences and columns for sample. This read count table can be used for
microbial profile comparison among different samples and the sequences provided in the table can be used to taxonomy assignment.
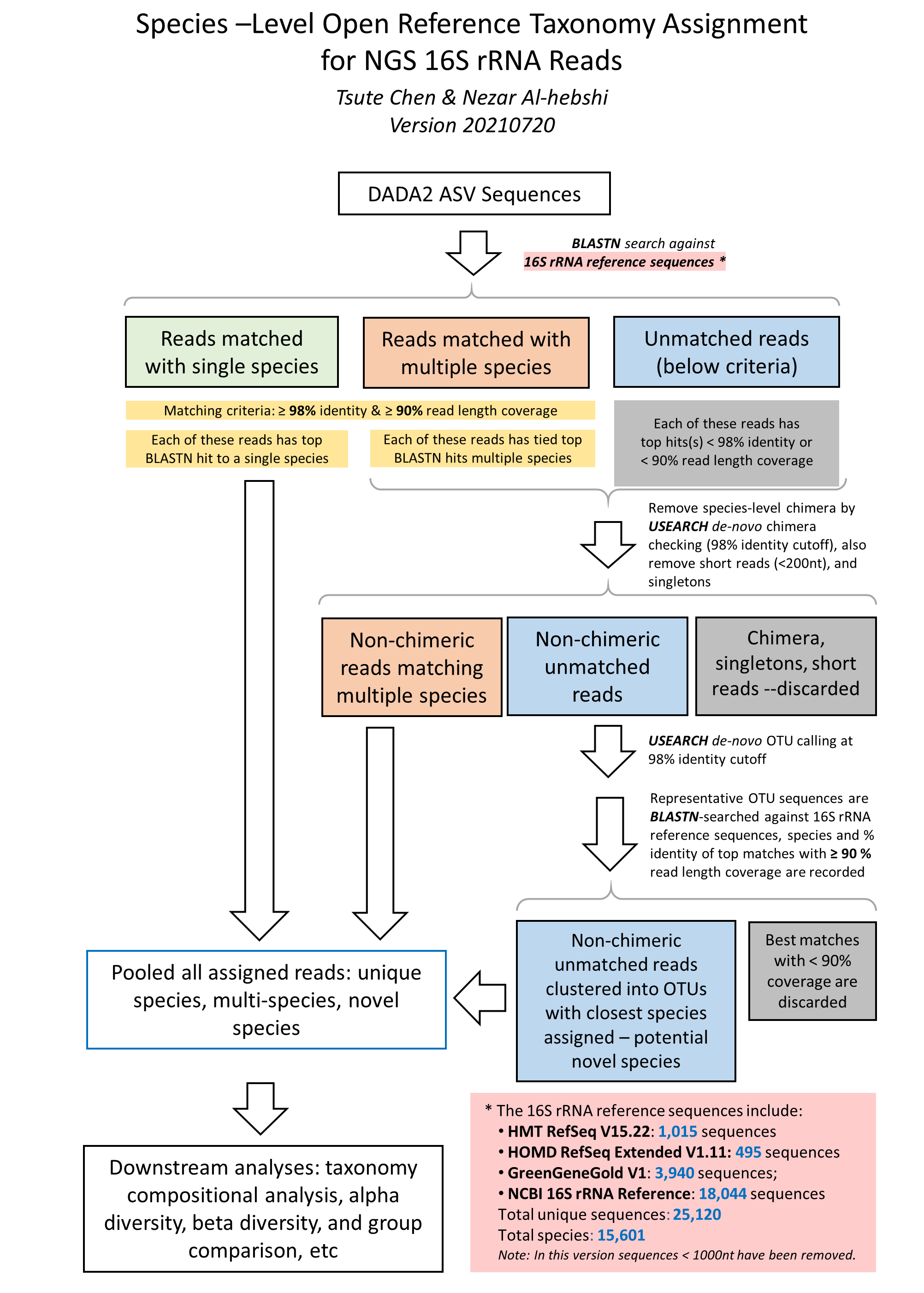
The species-level, open-reference 16S rRNA NGS reads taxonomy assignment pipeline
Version 20210310a
The close-reference taxonomy assignment of the ASV sequences using BLASTN is based on the algorithm published by Al-Hebshi et. al. (2015)[2].
1. Raw sequences reads in FASTA format were BLASTN-searched against a combined set of 16S rRNA reference sequences - the FOMC 16S rRNA Reference Sequences version 20221029 (https://microbiome.forsyth.org/ftp/refseq/).
This set consists of the HOMD (version 15.22 http://www.homd.org/index.php?name=seqDownload&file&type=R ), Mouse Oral Microbiome Database (MOMD version 5.1 https://momd.org/ftp/16S_rRNA_refseq/MOMD_16S_rRNA_RefSeq/V5.1/),
and the NCBI 16S rRNA reference sequence set (https://ftp.ncbi.nlm.nih.gov/blast/db/16S_ribosomal_RNA.tar.gz).
These sequences were screened and combined to remove short sequences (<1000nt), chimera, duplicated and sub-sequences,
as well as sequences with poor taxonomy annotation (e.g., without species information).
This process resulted in 1,015 full-length 16S rRNA sequences from HOMD V15.22, 356 from MOMD V5.1, and 22,126 from NCBI, a total of 23,497 sequences.
Altogether these sequence represent a total of 17,035 oral and non-oral microbial species.
The NCBI BLASTN version 2.7.1+ (Zhang et al, 2000) [3] was used with the default parameters.
Reads with ≥ 98% sequence identity to the matched reference and ≥ 90% alignment length
(i.e., ≥ 90% of the read length that was aligned to the reference and was used to calculate
the sequence percent identity) were classified based on the taxonomy of the reference sequence
with highest sequence identity. If a read matched with reference sequences representing
more than one species with equal percent identity and alignment length, it was subject
to chimera checking with USEARCH program version v8.1.1861 (Edgar 2010). Non-chimeric reads with multi-species
best hits were considered valid and were assigned with a unique species
notation (e.g., spp) denoting unresolvable multiple species.
2. Unassigned reads (i.e., reads with < 98% identity or < 90% alignment length) were pooled together and reads < 200 bases were
removed. The remaining reads were subject to the de novo
operational taxonomy unit (OTU) calling and chimera checking using the USEARCH program version v8.1.1861 (Edgar 2010)[4].
The de novo OTU calling and chimera checking was done using 98% as the sequence identity cutoff, i.e., the species-level OTU.
The output of this step produced species-level de novo clustered OTUs with 98% identity.
Representative reads from each of the OTUs/species were then BLASTN-searched
against the same reference sequence set again to determine the closest species for
these potential novel species. These potential novel species were pooled together with the reads that were signed to specie-level in
the previous step, for down-stream analyses.
Reference:
Al-Hebshi NN, Nasher AT, Idris AM, Chen T. Robust species taxonomy assignment algorithm for 16S rRNA NGS reads: application
to oral carcinoma samples. J Oral Microbiol. 2015 Sep 29;7:28934. doi: 10.3402/jom.v7.28934. PMID: 26426306; PMCID: PMC4590409.
Zhang Z, Schwartz S, Wagner L, Miller W. A greedy algorithm for aligning DNA sequences. J Comput Biol. 2000 Feb-Apr;7(1-2):203-14. doi: 10.1089/10665270050081478. PMID: 10890397.
Edgar RC. Search and clustering orders of magnitude faster than BLAST.
Bioinformatics. 2010 Oct 1;26(19):2460-1. doi: 10.1093/bioinformatics/btq461. Epub 2010 Aug 12. PubMed PMID: 20709691.
3. Designations used in the taxonomy:
1) Taxonomy levels are indicated by these prefixes:
k__: domain/kingdom
p__: phylum
c__: class
o__: order
f__: family
g__: genus
s__: species
Example:
k__Bacteria;p__Firmicutes;c__Clostridia;o__Clostridiales;f__Lachnospiraceae;g__Blautia;s__faecis
2) Unique level identified – known species:
k__Bacteria;p__Firmicutes;c__Clostridia;o__Clostridiales;f__Lachnospiraceae;g__Roseburia;s__hominis
The above example shows some reads match to a single species (all levels are unique)
3) Non-unique level identified – known species:
k__Bacteria;p__Firmicutes;c__Clostridia;o__Clostridiales;f__Lachnospiraceae;g__Roseburia;s__multispecies_spp123_3
The above example “s__multispecies_spp123_3” indicates certain reads equally match to 3 species of the
genus Roseburia; the “spp123” is a temporally assigned species ID.
k__Bacteria;p__Firmicutes;c__Clostridia;o__Clostridiales;f__Lachnospiraceae;g__multigenus;s__multispecies_spp234_5
The above example indicates certain reads match equally to 5 different species, which belong to multiple genera.;
the “spp234” is a temporally assigned species ID.
4) Unique level identified – unknown species, potential novel species:
k__Bacteria;p__Firmicutes;c__Clostridia;o__Clostridiales;f__Lachnospiraceae;g__Roseburia;s__ hominis_nov_97%
The above example indicates that some reads have no match to any of the reference sequences with
sequence identity ≥ 98% and percent coverage (alignment length) ≥ 98% as well. However this groups
of reads (actually the representative read from a de novo OTU) has 96% percent identity to
Roseburia hominis, thus this is a potential novel species, closest to Roseburia hominis.
(But they are not the same species).
5) Multiple level identified – unknown species, potential novel species:
k__Bacteria;p__Firmicutes;c__Clostridia;o__Clostridiales;f__Lachnospiraceae;g__Roseburia;s__ multispecies_sppn123_3_nov_96%
The above example indicates that some reads have no match to any of the reference sequences
with sequence identity ≥ 98% and percent coverage (alignment length) ≥ 98% as well.
However this groups of reads (actually the representative read from a de novo OTU)
has 96% percent identity equally to 3 species in Roseburia. Thus this is no single
closest species, instead this group of reads match equally to multiple species at 96%.
Since they have passed chimera check so they represent a novel species. “sppn123” is a
temporary ID for this potential novel species.
4. The taxonomy assignment algorithm is illustrated in this flow char below:
Read Taxonomy Assignment - Result Summary *
Code
Category
MPC=0% (>=1 read)
MPC=0.01%(>=354 reads)
A
Total reads
3,557,922
3,557,922
B
Total assigned reads
3,543,040
3,543,040
C
Assigned reads in species with read count < MPC
0
29,211
D
Assigned reads in samples with read count < 500
0
0
E
Total samples
48
48
F
Samples with reads >= 500
48
48
G
Samples with reads < 500
0
0
H
Total assigned reads used for analysis (B-C-D)
3,543,040
3,513,829
I
Reads assigned to single species
3,345,211
3,330,912
J
Reads assigned to multiple species
71,825
69,892
K
Reads assigned to novel species
126,004
113,025
L
Total number of species
875
381
M
Number of single species
458
321
N
Number of multi-species
25
8
O
Number of novel species
392
52
P
Total unassigned reads
14,882
14,882
Q
Chimeric reads
148
148
R
Reads without BLASTN hits
2,067
2,067
S
Others: short, low quality, singletons, etc.
12,667
12,667
A=B+P=C+D+H+Q+R+S
E=F+G
B=C+D+H
H=I+J+K
L=M+N+O
P=Q+R+S
* MPC = Minimal percent (of all assigned reads) read count per species, species with read count < MPC were removed.
* Samples with reads < 500 were removed from downstream analyses.
* The assignment result from MPC=0.1% was used in the downstream analyses.
Read Taxonomy Assignment - ASV Species-Level Read Counts Table
This table shows the read counts for each sample (columns) and each species identified based on the ASV sequences.
The downstream analyses were based on this table.
The species listed in the table has full taxonomy and a dynamically assigned species ID specific to this report.
When some reads match with the reference sequences of more than one species equally (i.e., same percent identiy and alignmnet coverage),
they can't be assigned to a particular species. Instead, they are assigned to multiple species with the species notaton
"s__multispecies_spp2_2". In this notation, spp2 is the dynamic ID assigned to these reads that hit multiple sequences and the "_2"
at the end of the notation means there are two species in the spp2.
You can look up which species are included in the multi-species assignment, in this table below:
Another type of notation is "s__multispecies_sppn2_2", in which the "n" in the sppn2 means it's a potential novel species because all the reads in this species
have < 98% idenity to any of the reference sequences. They were grouped together based on de novo OTU clustering at 98% identity cutoff. And then
a representative sequence was chosed to BLASTN search against the reference database to find the closest match (but will still be < 98%). This representative
sequence also matched equally to more than one species, hence the "spp" was given in the label.
In ecology, alpha diversity (α-diversity) is the mean species diversity in sites or habitats at a local scale.
The term was introduced by R. H. Whittaker[5][6] together with the terms beta diversity (β-diversity)
and gamma diversity (γ-diversity). Whittaker's idea was that the total species diversity in a landscape
(gamma diversity) is determined by two different things, the mean species diversity in sites or habitats
at a more local scale (alpha diversity) and the differentiation among those habitats (beta diversity).
Diversity measures are affected by the sampling depth. Rarefaction is a technique to assess species richness from the results of sampling. Rarefaction allows
the calculation of species richness for a given number of individual samples, based on the construction
of so-called rarefaction curves. This curve is a plot of the number of species as a function of the
number of samples. Rarefaction curves generally grow rapidly at first, as the most common species are found,
but the curves plateau as only the rarest species remain to be sampled [7].
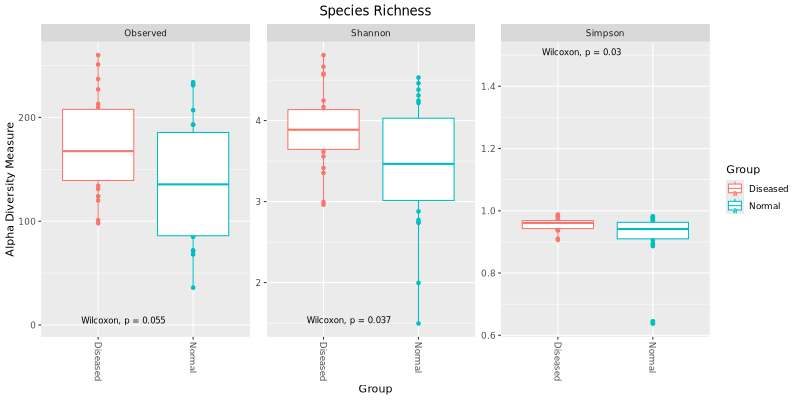
The two main factors taken into account when measuring diversity are richness and evenness.
Richness is a measure of the number of different kinds of organisms present in a particular area.
Evenness compares the similarity of the population size of each of the species present. There are
many different ways to measure the richness and evenness. These measurements are called "estimators" or "indices".
Below is a diversity of 3 commonly used indices showing the values for all the samples (dots) and in groups (boxes).
Alpha Diversity Box Plots for All Groups
Alpha Diversity Box Plots for Individual Comparisons at Species level
To test whether the alpha diversity among different comparison groups are different statistically, we use the Kruskal Wallis H test
provided the "alpha-group-significance" fucntion in the QIIME 2 "diversity" package. Kruskal Wallis H test is the non-parametric alternative
to the One Way ANOVA. Non-parametric means that the test doesn’t assume your data comes from a particular distribution. The H test is used
when the assumptions for ANOVA aren’t met (like the assumption of normality). It is sometimes called the one-way ANOVA on ranks,
as the ranks of the data values are used in the test rather than the actual data points. The H test determines whether the medians of two
or more groups are different.
Below are the Kruskal Wallis H test results for each comparison based on three different alpha diversity measures: 1) Observed species (features),
2) Shannon index, and 3) Simpson index.
Beta diversity compares the similarity (or dissimilarity) of microbial profiles between different
groups of samples. There are many different similarity/dissimilarity metrics [8].
In general, they can be quantitative (using sequence abundance, e.g., Bray-Curtis or weighted UniFrac)
or binary (considering only presence-absence of sequences, e.g., binary Jaccard or unweighted UniFrac).
They can be even based on phylogeny (e.g., UniFrac metrics) or not (non-UniFrac metrics, such as Bray-Curtis, etc.).
For microbiome studies, species profiles of samples can be compared with the Bray-Curtis dissimilarity,
which is based on the count data type. The pair-wise Bray-Curtis dissimilarity matrix of all samples can then be
subject to either multi-dimensional scaling (MDS, also known as PCoA) or non-metric MDS (NMDS).
MDS/PCoA is a
scaling or ordination method that starts with a matrix of similarities or dissimilarities
between a set of samples and aims to produce a low-dimensional graphical plot of the data
in such a way that distances between points in the plot are close to original dissimilarities.
NMDS is similar to MDS, however it does not use the dissimilarities data, instead it converts them into
the ranks and use these ranks in the calculation.
In our beta diversity analysis, Bray-Curtis dissimilarity matrix was first calculated and then plotted by the PCoA and
NMDS separately. Below are beta diveristy results for all groups together:
The above PCoA and NMDS plots are based on count data. The count data can also be transformed into centered log ratio (CLR)
for each species. The CLR data is no longer count data and cannot be used in Bray-Curtis dissimilarity calculation. Instead
CLR can be compared with Euclidean distances. When CLR data are compared by Euclidean distance, the distance is also called
Aitchison distance.
Below are the NMDS and PCoA plots of the Aitchison distances of the samples:
NMDS and PCoA Plots for Individual Comparisons at Species level
Interactive 3D PCoA Plots - Bray-Curtis Dissimilarity
Interactive 3D PCoA Plots - Euclidean Distance
Interactive 3D PCoA Plots - Correlation Coefficients
Group Significance of Beta-diversity Indices
To test whether the between-group dissimilarities are significantly greater than the within-group dissimilarities,
the "beta-group-significance" function provided in the QIIME 2 "diversity" package was used with PERMANOVA
(permutational multivariate analysis of variance) as the group significant testing method.
Three beta diversity matrics were used: 1) Bray–Curtis dissimilarity 2) Correlation coefficient matrix , and 3) Aitchison distance
(Euclidean distance between clr-transformed compositions).
16S rRNA next generation sequencing (NGS) generates a fixed number of reads that reflect the proportion of different
species in a sample, i.e., the relative abundance of species, instead of the absolute abundance.
In Mathematics, measurements involving probabilities, proportions, percentages, and ppm can all
be thought of as compositional data. This makes the microbiome read count data “compositional”
(Gloor et al, 2017). In general, compositional data represent parts of a whole which only
carry relative information [9].
The problem of microbiome data being compositional arises when comparing two groups of samples for
identifying “differentially abundant” species. A species with the same absolute abundance between two
conditions, its relative abundances in the two conditions (e.g., percent abundance) can become different
if the relative abundance of other species change greatly. This problem can lead to incorrect conclusion
in terms of differential abundance for microbial species in the samples.
When studying differential abundance (DA), the current better approach is to transform the read count
data into log ratio data. The ratios are calculated between read counts of all species in a sample to
a “reference” count (e.g., mean read count of the sample). The log ratio data allow the detection of DA
species without being affected by percentage bias mentioned above
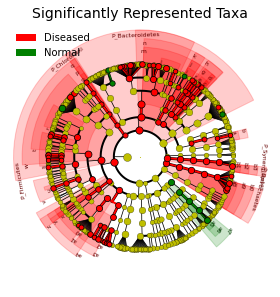
In this report, a compositional DA analysis tool “ANCOM” (analysis of composition of microbiomes)
was used [10]. ANCOM transforms the count data into log-ratios and thus is more suitable for comparing
the composition of microbiomes in two or more populations. "ANCOM" generates a table of features with
W-statistics and whether the null hypothesis is rejected. The “W” is the W-statistic, or number of
features that a single feature is tested to be significantly different against. Hence the higher the "W"
the more statistical sifgnificant that a feature/species is differentially abundant.
References:
Gloor GB, Macklaim JM, Pawlowsky-Glahn V, Egozcue JJ. Microbiome Datasets Are Compositional: And This Is Not Optional. Front Microbiol.
2017 Nov 15;8:2224. doi: 10.3389/fmicb.2017.02224. PMID: 29187837; PMCID: PMC5695134.
Mandal S, Van Treuren W, White RA, Eggesbø M, Knight R, Peddada SD. Analysis of composition of
microbiomes: a novel method for studying microbial composition. Microb Ecol Health Dis.
2015 May 29;26:27663. doi: 10.3402/mehd.v26.27663. PMID: 26028277; PMCID: PMC4450248.
Starting with version V1.2, we include the results of ANCOM-BC (Analysis of Compositions of
Microbiomes with Bias Correction) (Lin and Peddada 2020) [11]. ANCOM-BC is an updated version of "ANCOM" that:
(a) provides statistically valid test with appropriate p-values,
(b) provides confidence intervals for differential abundance of each taxon,
(c) controls the False Discovery Rate (FDR),
(d) maintains adequate power, and
(e) is computationally simple to implement.
The bias correction (BC) addresses a challenging problem of the bias introduced by differences in
the sampling fractions across samples. This bias has been a major hurdle in performing DA analysis of microbiome data.
ANCOM-BC estimates the unknown sampling fractions and corrects the bias induced by their differences among samples.
The absolute abundance data are modeled using a linear regression framework.
Starting with version V1.43, ANCOM-BC2 is used instead of ANCOM-BC, So that multiple pairwise directional test can be performed (if there are more than two gorups in a comparison).
When performing pairwise directional test, the mixed directional false discover rate (mdFDR) is taken into account. The mdFDR
is the combination of false discovery rate due to multiple testing, multiple pairwise comparisons, and directional tests within
each pairwise comparison. The mdFDR is adopted from (Guo, Sarkar, and Peddada 2010 [12]; Grandhi, Guo, and Peddada 2016 [13]). For more detail
explanation and additional features of ANCOM-BC2 please see author's documentation.
References:
Lin H, Peddada SD. Analysis of compositions of microbiomes with bias correction.
Nat Commun. 2020 Jul 14;11(1):3514. doi: 10.1038/s41467-020-17041-7.
PMID: 32665548; PMCID: PMC7360769.
Guo W, Sarkar SK, Peddada SD. Controlling false discoveries in multidimensional directional decisions, with applications to gene expression data on ordered categories. Biometrics. 2010 Jun;66(2):485-92. doi: 10.1111/j.1541-0420.2009.01292.x. Epub 2009 Jul 23. PMID: 19645703; PMCID: PMC2895927.
Grandhi A, Guo W, Peddada SD. A multiple testing procedure for multi-dimensional pairwise comparisons with application to gene expression studies. BMC Bioinformatics. 2016 Feb 25;17:104. doi: 10.1186/s12859-016-0937-5. PMID: 26917217; PMCID: PMC4768411.
"ALDEx2 is a compositional data analysis tool designed to enhance the statistical analysis of high-throughput sequencing datasets,
including RNA-seq, ChIP-seq, 16S rRNA gene sequencing, metagenomic analysis, and selective growth experiments.
Despite the fundamental similarities in data structure across these various experimental designs—namely,
counts of sequencing reads mapped to numerous features—traditional data analysis methods
have remained disparate and non-transferable between experiment types.
ALDEx2 addresses this challenge by employing compositional data analysis methods from the physical and geological sciences,
which convert raw data into relative abundances. This transformation leads to analyses that are more robust and reproducible.
Utilizing Bayesian methods to infer technical and statistical errors, ALDEx2 has demonstrated its applicability and effectiveness
across diverse datasets. It accurately identifies differential abundance and the direction of changes in selective growth experiments,
aligns closely with leading tools in identifying differentially expressed genes in RNA-seq datasets,
and successfully distinguishes differential taxa in the Human Microbiome Project 16S rRNA gene abundance dataset."
In this paired-sample differential abundance test, ALDEx2 was used with the Wilcoxon rank-sum test to identify features at different taxonomy ranks (from Phylum to Species)
that are significantly differentially abundant between two conditions. p-values were adjusted using "Holm" or "Benjamini-Hochberg" (BH) method to control the false discovery rate (FDR).
The simplest but strict p-value adjustment method is the Bonferroni method in which the p-values are multiplied by the number of comparisons.
Both Holm (1979) and Benjamini & Hochberg (1995) ("BH" or its alias "fdr") provide less conservative corrections.
In the below ALDEx2 result folder, comparisons were done with these two adjustment methods. Also, analyses were done with and without "paired sample" options for comparison.
References:
Fernandes AD, Macklaim JM, Linn TG, Reid G, Gloor GB. ANOVA-like differential expression (ALDEx) analysis for mixed population RNA-Seq. PLoS One. 2013 Jul 2;8(7):e67019. doi: 10.1371/journal.pone.0067019. PMID: 23843979; PMCID: PMC3699591.
Fernandes AD, Reid JN, Macklaim JM, McMurrough TA, Edgell DR, Gloor GB. Unifying the analysis of high-throughput sequencing datasets: characterizing RNA-seq, 16S rRNA gene sequencing and selective growth experiments by compositional data analysis. Microbiome. 2014 May 5;2:15. doi: 10.1186/2049-2618-2-15. PMID: 24910773; PMCID: PMC4030730.
Bonferroni, C. E., Teoria statistica delle classi e calcolo delle probabilità, Pubblicazioni del R Istituto Superiore di Scienze Economiche e Commerciali di Firenze 1936
Holm, S. (1979). A simple sequentially rejective multiple test procedure. Scandinavian Journal of Statistics, 6, 65--70. http://www.jstor.org/stable/4615733.
Benjamini, Y., and Hochberg, Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. Journal of the Royal Statistical Society Series B, 57, 289--300. http://www.jstor.org/stable/2346101.
LEfSe (Linear Discriminant Analysis Effect Size) is an alternative method to find "organisms, genes, or
pathways that consistently explain the differences between two or more microbial communities" (Segata et al., 2011) [19].
Specifically, LEfSe uses rank-based Kruskal-Wallis (KW) sum-rank test to detect features with significant
differential (relative) abundance with respect to the class of interest. Since it is rank-based, instead of proportional based,
the differential species identified among the comparison groups is less biased (than percent abundance based).
Reference:
Segata N, Izard J, Waldron L, Gevers D, Miropolsky L, Garrett WS, Huttenhower C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011 Jun 24;12(6):R60. doi: 10.1186/gb-2011-12-6-r60. PMID: 21702898; PMCID: PMC3218848.
To analyze the co-occurrence or co-exclusion between microbial species among different samples, network correlation
analysis tools are usually used for this purpose. However, microbiome count data are compositional. If count data are normalized to the total number of counts in the
sample, the data become not independent and traditional statistical metrics (e.g., correlation) for the detection
of specie-species relationships can lead to spurious results. In addition, sequencing-based studies typically
measure hundreds of OTUs (species) on few samples; thus, inference of OTU-OTU association networks is severely
under-powered. Here we use SPIEC-EASI (SParse InversECovariance Estimation
for Ecological Association Inference), a statistical method for the inference of microbial
ecological networks from amplicon sequencing datasets that addresses both of these issues (Kurtz et al., 2015) [20].
SPIEC-EASI combines data transformations developed for compositional data analysis with a graphical model
inference framework that assumes the underlying ecological association network is sparse. SPIEC-EASI provides
two algorithms for network inferencing – 1) Meinshausen-Bühlmann's neighborhood selection (MB method) and inverse covariance selection
(GLASSO method, i.e., graphical least absolute shrinkage and selection operator). This is fundamentally distinct from SparCC, which essentially estimate pairwise correlations. In addition
to these two methods, we provide the results of a third method - SparCC (Sparse Correlations for Compositional Data)(Friedman & Alm 2012)[21], which
is also a method for inferring correlations from compositional data. SparCC estimates the linear Pearson correlations between
the log-transformed components.
References:
Kurtz ZD, Müller CL, Miraldi ER, Littman DR, Blaser MJ, Bonneau RA. Sparse and compositionally robust inference of microbial ecological networks. PLoS Comput Biol. 2015 May 7;11(5):e1004226. doi: 10.1371/journal.pcbi.1004226. PMID: 25950956; PMCID: PMC4423992.
The results of this analysis are for research purpose only. They are not intended to diagnose, treat, cure, or prevent any disease. Forsyth and FOMC
are not responsible for use of information provided in this report outside the research area.













{kind=link}
{kind=link}
{kind=link}